| هسته محل ذخیره اطلاعات ژنتیکی و مرکز کنترل سلول یوکاریوتی است. محتویات هسته در یوکاریوتها توسط غشای هسته احاطه شده است و اندامکی به نام هسته را بوجود آورده است. ولی چون سلولهای پروکاریوتی فاقد غشای هسته هستند بدون هسته محسوب میشوند. |
اطلاعات اولیه
هسته در سال 1831 توسط Robert Brown در سلولهای اپیدرمی ثعلبیان کشف شد و به عنوان بخشی متراکم ، پایدار و موجود در همه سلولها در نظر گرفته شد. هسته یک ساختار فشرده با پیچ و تابهای زیادی را داراست و با پروتئین همراه میباشد. به چنین مجموعه فشردهای همراه با پروتئین ، کروماتین میگویند. هسته واجد غشای دو لایهای موسوم به پوشش هستهای است و در این پوشش حفرهها یا روزنههایی موسوم به منفذ پیچیده هستهای است که از طریق آنها عمل تبادل مواد بین هسته و سیتوپلاسم انجام میگیرد. برای بررسی ساختمان عمومی هسته میتوان از میکروسکوپ نوری و میکروسکوپ الکترونی استفاده کرد.
| عکس پیدا نشد |
شکل و محل و تعداد هسته در سلولها
هسته در بیشتر سلولها کروی یا بیضوی است. در سلولهای پارانشیمی بالغ گیاهان عدسی شکل ، در سلولهای عضلانی مخطط جانوران و سلولهای پروکامبیومی گیاهان استوانهای شکل ، در سلولهای آبکش بالغ و سلولهای انگل زده چند بخشی است. در عدهای از سلولها هسته چند بخشی است مثل گویچههای سفید خون چند هستهای و یا سلولهای استئوکلاست (استخوان خوار). در بیشتر سلولها هسته در مرکز قرار دارد. در سلولهای گیاهی به علت رشد واکوئلها ، هسته در کنار غشا قرار میگیرد و در سلولهای عضلانی مخطط هسته در بخشهای کناری قرار دارد.
در جلبک استابولاریا هسته در بخش ریزوئیدی (ریشه نما) یا مجاور با آن قرار دارد. اغلب سلولها دارای یک هسته هستند. با وجود این برخی جانداران ابتدایی و یا سلولهای جانداران پیشرفته بیش از یک هسته دارند. برای مثال حدود 20 درصد از سلولهای کبدی و یا عده زیادی از سلولهای ریسه قارچها دو هستهای هستند. سلولهای عضلانی مخطط ساختمان سنوسیتی دارند یعنی در یک سیتوپلاسم مشترک چندین هسته پراکنده است. این سلولها ابتدا یک هستهای بودهاند که به دلیل تقسیمات مکرر هسته بدون آنکه سیتوپلاسم تقسیم شود به حالت سنوسیتی درآمدهاند.
| نسبت حجم هسته به حجم سیتوپلاسم را نسبت نوکلئوپلاسمی میگویند. این نسبت برای سلولهایی که در یک مرحله رشد و در شرایط مشابه باشند ثابت است. |
پوشش هستهای
اطراف هسته سلولهای یوکاریوتی را پوشش هستهای شامل غشای بیرونی ، غشای درونی ، فضای بین دو غشا و منافذ هستهای پوشانیده است.
غشای بیرونی
از دو لایه فسفولیپیدی و پروتئینهای پراکنده در بین آنها تشکیل شده است که شباهت زیادی به غشای شبکه آندوپلاسمی دارد و در سطح آن ریبوزومها قرار گرفتهاند. بخشهایی از غشای بیرونی با شبکه آندوپلاسمی دانهدار ، پیوستگی دارد و منشا غشای هسته نیز از شبکه آندوپلاسمی دانهدار است.
فضای بین دو غشا
فضای بین دو غشای یا فضای دور هستهای فضایی به وسعت 60 تا 100 آنگستروم است که وسعت آن در همه جای پوشش هستهای یکنواخت نیست. در برخی نواحی وسیعتر و در محل منافذ یا سوراخهای هستهای که دو غشا پوشش هستهای بهم میرسند وسعت فضای دور هستهای به صفر میرسد.
غشای داخلی
غشایی زیستی به ضخامت حدود 60 تا 70 آنگستروم ، شبیه غشای شبکه آندوپلاسمی و فاقد ریبوزوم است. غشای داخلی با واسطه پروتئینهای لامینایی با کروماتین ارتباط دارد.
| عکس پیدا نشد |
منافذ غشای هسته
در پوشش هستهای ساختمانهای پروتئینی فعال و ویژهای به اسم منافذ هستهای وجود دارد. وجود این منافذ بوسیله هرتویگ در سال 1876 برای اولین بار پیشبینی شد. قطر منافذ به اندازهای است که به مولکولهای پروتئین ، انواع RNAها و حتی زیر واحدهای ریبوزومی اجازه عبور میدهد. پروتئینهای سیتوپلاسمی که وارد هسته میشوند از جمله پروتئینهای هیستونی دارای یک بخش نشانه هستند که به کمک آن از بازگشت آنها به سیتوپلاسم جلوگیری میشود. منافذ هستهای عبور یونهای منفی را تسهیل میکنند.
منافذ هستهای ساختمانهای دائمی و پایدار نیستند و متناسب با نیاز سلول ایجاد یا ناپدید میشوند. در سلولهای با فعالیت متابولیکی بالا که مبادله مواد بین هسته و سیتوپلاسم زیاد است تعداد منافذ هسته نیز زیاد است و در سلولهایی که تبادلات هسته و سیتوپلاسم کم است تعداد منافذ کاهش مییابد. هر منفذ بوسیله مجموعهای از ذرات متراکم احاطه شده است. این ساختمانهای پروتئینی را بر روی هم مجموعه منفذی یا منفذ پیچیده هستهای مینامند که شامل بخشهای زیر است.
- یک حلقه یا آنولوس که از 8 پروتئین گرانولی کناری تشکیل شده است و در سطح سیتوزولی قرار دارد.
- یک حلقه یا آنولوس که این هم از 8 پروتئین گرانولی کناری تشکیل شده است و در سطح نوکلئوپلاسمی قرار دارد.
- کانال مرکزی یا درپوش که این کانال محل عبور مواد میباشد و در مرکز منفذ قرار دارد.
- منفذ از ترکیباتی به نام Annular Material پر شده و به سمت نوکلئوپلاسم (شیره هسته) و سیتوپلاسم بیرون زده است.
شیره هسته ( نوکلئوپلاسم یا کاریولنف )
شیره هسته مایعی است که درون هسته را پر کرده است و از نظر کلی شبیه سیتوزول و کمی متراکمتر از آن با PH اسیدی است. شیره هسته یا ماتریکس هسته حاوی آب 10 درصد از کل پروتئینهای هستهای ، 30 درصد از کل RNA و 2 تا 5 درصد از کل فسفولیپیدهای هستهای را شامل میشود. مقدار کمی لیپید و نیز مقداری گلوسیدهای (قندها) موثر در تشکیل نوکلئوتید مثل ریبوز و دزوکسی ریبوز در آن وجود دارند.
یونهای موجود در شیره هسته
در شیره هسته یونهای 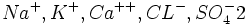 وجود دارد. یونهای
وجود دارد. یونهای 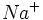 موجود در هسته ده برابر بیشتر از یونهای موجود در سیتوپلاسم میباشد. و از این لحاظ مساوی محیط خارج سلول است. یون در هسته برای نگهداری ساختمان مولکول DNA لازم میباشد. یون
موجود در هسته ده برابر بیشتر از یونهای موجود در سیتوپلاسم میباشد. و از این لحاظ مساوی محیط خارج سلول است. یون در هسته برای نگهداری ساختمان مولکول DNA لازم میباشد. یون 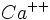 مانع تجمع هیستونها بر روی DNA و اثری در جهت فعال کردن تنظیم بیان ژن و افزایش نسخه برداری دارد.
مانع تجمع هیستونها بر روی DNA و اثری در جهت فعال کردن تنظیم بیان ژن و افزایش نسخه برداری دارد.
پروتئینهای شیره هسته
این پروتئینها به دو گروه ساختمانی شامل هیستونها و تنظیمی شامل انواع غیر هیستونی و آنزیمی تقسیم میشوند.
- پروتامینها : از مهمترین پروتئینهای موجود درشیره هسته سلولهای جنسی و زایشی هستند که خاصیت قلیایی دارند.
- پروتئینهای هیستونی : با دارا بودن خاصیت قلیایی به DNA متصل شده و نوکلئوزومها یا واحدهای تکراری DNA را میسازند.
- پروتئینهای غیر هیستونی : دارای خاصیت اسیدی هستند.
- _پروتئینهای آنزیمی :__ پروتئینهای آنزیمی شیره هسته شامل DNA پلیمرازها ، RNA پلیمرازها ، لیگازها ، DNase ، RNase ، GTPase ، ATPase ، نوکلئوزید فسفریلاز است.
ساختمان شیره هسته
شیره هسته یا نوکلئوپلاسم حاوی انواع گوناگونی از گرانولهای بین کروماتینی و گرانولهای اطراف کروماتینی است.
- گرانولهای بین کروماتینی ( ICG ) : ذراتی است به قطر 20 تا 25 نانومتر که توسط رشتههایی به یکدیگر متصل شدهاند و به فرم دستجاتی در فضای بین کروماتین اکثر هستهها دیده میشود.
- گرانولهای اطراف کروماتینی ( PCG ): در اطراف هتروکروماتین متراکم قرار داشته و مانند گرانولهای منفرد به نظر میآیند. این گرانولها به قطر 30 تا 50 نانومتر بوده و توسط گرانولهایی به قطر 25 نانومتر احاطه شدهاند. این گرانولها در بسیاری از هسته سلولها دیده شده و از تراکم فیبرهای بسته بندی شده به قطر 3 نانومتر تشکیل شدهاند. این گرانولها از RNA دارای وزن مولکولی کم و ضریب رسوب 4.7s و حداقل 2 پروتئین تشکیل شده است.
اسکلت هستهای
مجموعه منفذی شبکه لامینایی به علاوه اسکلت هستهای درونی را روی هم اسکلت هستهای گویند.
شبکه لامینایی
شبکه لامینائی یا لامینها پروتئینهای خاصی هستند که تریمرهایی تشکیل میدهند متشکل از سه مونومر که با A ، B ، C یا a ، b ، c معرفی میشوند. تریمرهای لامینایی که از سه مونومر A ، B و C تشکیل یافتهاند همانند شبکه تورینهای بهم میپیوندند و اسکلت هستهای را میسازند. بخش محیطی (اطرافی) اسکلت هستهای تراکم بیشتری دارد و بخش درونی آن کم تراکمتر و دارای حالت اسفنجی است. شبکه لامینایی را اغلب شبکه بسیار ظریفی میدانند که در مقابل سطح درونی پوشش هستهای قرار گرفتهاند و با مجموعههای منفذی اتصالهایی دارد. شبکه لامینایی ساختمانی ظریف اما چسبنده دارد که پس از حذف پوشش هستهای و حل کردن پروتئین و DNAی موجود در کروماتین بصورت پاکت نازکی باقی میماند و اندازه و حالت هسته را حفظ میکند.
لامین نقش ساختمانی مهمی در تعیین شکل و وضعیت هسته بازی میکند. لامینها مولکولهای بسیار پایداری هستند. به حسب وضع لامینها در طول تقسیم سلولی دو نوع لامین در نظر میگیرند. لامینایی که همیشه چسبیده به پوشش هستهای باقی میماند (لامین B در پستانداران) و آنهایی که هنگام تقسیم در سیتوپلاسم حل میشوند (لامین A و C در پستانداران). لامینها به اندازه کافی بخشهای آبگریز برای نفوذ به دو لایه لیپیدی غشای داخل هسته را ندارند اما با واسطه یک نوع از پروتئینهای درون غشایی به آن متصلند.
اسکلت هستهای درونی
هنگامی که هستهها جدا شده از سلول بوسیله نوکلئازها از جمله DNase و RNase که به ترتیب DNA و RNA را هضم میکنند یا با محلولهای نمکی قوی که پروتئینهای کروی را حل میکنند، تیمار شوند یک شبکه سه بعدی درون هستهای باقی میماند که با میکروسکوپ الکترونی قابل مشاهده است و در مجموعه شیره هسته پراکنده است. این ساختمان را اسکلت درونی هستهای میگویند. به نظر میرسد که این شبکه برای سازمانیابی کروماتین مهم است. اسکلت هستهای درونی از دو شبکه روی هم تشکیل شده است که عبارتند از:
- شبکه اول از پروتئینهای رشتهای ساخته شده و نقش پشتیبان را برای شبکه دوم دارد.
- شبکه دوم از پروتئینهای گویچهای یا کروی ساخته شده که بین آنها پروتئینهای آنزیمی دخالت کننده در رونویسی و همانند سازی نیز در بین آنها وجود دارد.
هستک
هستک در هسته انترفاز دیده میشود از مرحله پروفاز تا اوایل تلوفاز دیده نمیشود. هستک یک اندامک درون هستهای و بدون غشا در شیره هسته میباشد. تعداد معمول آن یک یا دو عدد در هسته هر سلول ، گاهی چند عدد و در هسته اووسیت دوزیستان تعداد زیادی هستک وجود دارد هستکها اغلب کروی شکل و به صورت ذرات متراکم هستند.
بین درشتی هستک و فعالیت بیوسنتزی پروتئینی سلول وابستگی وجود دارد. هرچه سلولها دارای سنتز پروتئین بیشتر باشند هستکهای درشتتری دارند مثل اووسیتها ، سلولهای ترشحی و نورونها. در مقابل در اسپرماتیدها و سلولهای عضلانی هستکها کوچکترند. هستک حاوی RNA فراوان است و یک منطقه کروماتینی متراکم و کم و بیش حلقهدار اطراف هستک را احاطه میکند هستک جایگاه تشکیل ریبوزوم است.
| عکس پیدا نشد |
کروماتین
ترکیب اصلی هسته ، کروماتین است شبکه کروماتینی از درهم رفتن رشتههای کروماتینی تشکیل شده و این رشتهها در حقیقت حالت بسیار کم تراکم شدهای از کروموزومها هستند. رشد کروماتین یا DNAی انترفازی ، مجموعه مولکولی پیچیدهای است که در آن DNA ، دارای اطلاعات ژنتیکی و پروتئینهای مختلف وابسته به آن نقش ساختمانی یا عملی و نیز مقداری از RNAها وجود دارد.
مباحث مرتبط با عنوان
- پروتئین
- ساختمان DNA
- سلول انسانی
- سلول گیاهی
- سیتوپلاسم
- شبکه آندوپلاسمی
- ریبوزوم
- غشای سلولی
- کروموزوم
- میتوز
نظرات شما عزیزان:
.: Weblog Themes By Pichak :.